Reporting podľa článku 17 smernice o biotopoch za obodbie rokov 2007 - 2012 cela správa [pdf]
Úvod
Štátna ochrana prírody Slovenskej republiky v spolupráci s ďalšími organizáciami a expertmi v závere roka 2013 pripravila oficiálnu správu o stave biotopov a druhov európskeho významu za obdobie rokov 2007 – 2012 a prostredníctvom odobrenia zo strany Ministerstva životného prostredia Slovenskej republiky správu odovzdala Európskej komisii. Správa poskytuje detailné informácie o rozšírení, areáli, vplyvoch a ohrozeniach, hodnotení stavu a ďalších parametroch pre každý biotop/druh eur. významu zvlášť a teda poskytuje najaktuálnejšie a najkomplexnejšie vecné informácie o uvedenej problematike. Pripravený dokument v úvodnej časti pojednáva o procese a metodike prípravy samotnej správy. V ďalšej časti obsahuje sumárne vyhodnotenia, štatistiky, grafy, ktoré analyzujú samotné výsledky správy. Nasledovnú a najrobustnejšiu časť potom tvoria samotné „reporty” pre jednotlivé biotopy a druhy, v ktorých sú uvedené čiastkové informácie z databázy ako aj mapy rozšírenia a mapy areálu pre jednotlivé biotopy a druhy eur.významu.
1 Informácie o procese prípravy správy
1.1 Použité údaje pri príprave správy (reportingu)
V prípade vyšších rastlín sú pre reporting použité najmä údaje z Čiastkového monitorovacieho systému BIOTA (ČMS Biota) a projektu „ŠF Monitoring". Od roku 2005 podlieha systematickému monitoringu 37 rastlinných druhov európskeho významu. Monitorovaná je väčšina známych lokalít jednotlivých druhov, u vzácnych druhov sú monitorované všetky ich známe lokality. Kvalita údajov je teda pri vyšších rastlinách založená na podrobnom monitoringu, ktorý kontinuálne realizujú pracovníci ŠOP SR. Spracovaný reporting stavu živočíchov vychádza najmä z údajov získaných z Informačného systému taxónov a biotopov (ISTB) - výskytové dáta, ČMS Biota (druhy Emys orbicularis, Lutra lutra, Rupicapra rupicapra tatrica, Marmota marmota latirostris, Spermophilus citellus), z publikovaných údajov, zadaných výskumov (ryby, chrobáky, vážky a motýle v rokoch 2006 - 2008) a od expertov pre jednotlivé skupiny.
Kvalita údajov o biotopoch do veľkej miery závisí od údajov zapracovaných ďalšími organizáciami (DAPHNE, Národné lesnícke centrum, atď.), čiastkovo sú údaje spracované z niektorých projektov vykonávanými ŠOP SR (napr. mokrade, modráčiky). Systematický monitoring všetkých druhov a biotopov európskeho významu začína až rokom 2013 prostredníctvom projektu „ŠF Monitoring". Údaje z tohto roku monitorovania sú väčšinou použité len na spresnenie informácií a budú slúžiť predovšetkým ako podklad pre prípravu reportu za obdobie 2013 - 2018.
Dôležitým podkladom pre prípravu uvedenej správy za roky 2007 - 2012 o stave druhov a biotopov je prvotný reporting (2004-2006), ktorý definoval výskyt, areál, trendy, stav druhov a biotopov a ďalšie stanovené parametre, ktoré terajší reporting aktualizuje a dopĺňa. Upozorňujeme, že použité dáta pre prípravu reportingu boli veľmi obmedzené a vzhľadom na doteraz chýbajúci celoplošný monitoring má tomu primeranú kvalitu aj samotná výsledná správa. Neexistencia komplexného mapovania biotopov a monitoringu za posledné obdobie teda vo výraznej miere ovplyvňuje finálny výsledok, pretože pre niektoré polia a ich adekvátne vyplnenie neexistujú dostatočné podklady, na čo môže EK upozorniť.
1.2 Postup pri spracovávaní jednotlivých údajov pre reporting
1.2.1 Úvodná fáza prípravy
Úvodná fáza prípravy správy vyžadovala predovšetkým predpripravenie dát v požadovanej štruktúre takým spôsobom, aby spĺňal základné požiadavky formátu a bolo možné ďalej s údajmi pracovať.
Vzhľadom na nový formát reportingu Európskej komisie, ktorý upravuje niektoré polia, dopĺňa a mení štruktúru samotnej databázy a definuje presne požadované štandardy na GIS vrstvy oproti pôvodnému formátu správy, bolo potrebné v prvom kroku pôvodné údaje modifikovať, doplniť a prispôsobiť novému formátu.
Transformácia a doplnenie databázy sa týkalo nasledovných skupín údajov:
1. Kompatibilné údaje, ktoré boli v súlade z novým formátom boli prevzaté do databázy „art 17 reporting tool" 1:1.
2. Upravené údaje si vyžadovali prispôsobiť formát prostredníctvom databázových krokov, tak aby mohli byť vložené do samotnej databázy „art 17 reporting tooľ' už v požadovanej upravenej verzii.
3. Transformované údaje, si vyžadovali úplnú zmenu. Týkali sa prekódovania vplyvov a ohrození podľa aktuálne platného číselníka, kde vo viacerých krokoch boli priradené nové kódy. Vplyvy a ohrozenia boli kompletne zrevidované pre všetky biotopy a druhy.
4. Chýbajúce údaje, ktoré v predošlom formáte neboli požadované, boli doplnené na základe dostupných podkladov.
V nasledujúcej časti uvádzame niektoré podrobnosti procesu transformácie a prípravy v úvodnej fáze.
Postup pri stanovení veľkosti populácií (v jedincoch) je jednou z nových súčastí terajšieho formátu. Z databázy informačného systému taxónov a biotopov (ISTB) boli vyexportovane všetky údaje o druhoch eur. významu. Dáta boli rozdelené zvlášť pre alpský a panónsky bioregión, pretože hodnotenie veľkosti populácie musí byť spracované samostatne. Následne boli prostredníctvom funkcie „dissolve" zlúčené všetky jednotlivé druhy s pridaním spočítania početnosti druhov v jednotlivých záznamoch. Vychádzali sme z predpokladu, že celoslovensky zbierané údaje prostredníctvom jednotlivých expertov na správach ŠOP SR a externých spolupracovníkov budú dostatočne štatisticky preukaznou vzorkou pre čiastkové údaje o populácii jednotlivých druhov. Následne boli údaje pre jednotlivé druhy extrapolované o pomernú časť, v ktorej sa zohľadnil fakt, či sa jedná o druh pomerne poznaný, stavovca alebo bezstavovca a a či je atraktívnym druhom, ktorému sa venuje viac pozornosti a teda existuje pre neho aj viac údajov. Následne bola stanovená početnosť zaradená do odporúčaného intervalu podľa manuálu pre reporting. Uvedený postup bol použitý len pri druhoch, pri ktorých neexistovali presnejšie údaje o početnosti. V prípade druhov rastlín boli použité presné rozsahy početnosti na základe podrobného monitoringu. V prípade niektorých lepšie poznaných druhov, niektorých cicavcov (napr. kamzík, syseľ, vydra) boli taktiež použité presnejšie čísla, keďže tieto druhy sú lepšie poznané a teda aj ich početnosť je presnejšie stanovená.
Stanovenie početnosti v územiach Nátura 2000 bolo následne spracované na základe údajov v SDF nasledovným spôsobom. V prvom kroku boli územia rozdelené podľa bioregiónov na alpský a panónsky. Následne boli vyexportovane jednotlivé hodnotenia populácie podľa samotných území a zadefinovaný priemer pre jednotlivé hodnotenia A, B, C, D v percentách. Priemerný rozsah percent, ktorý vyšiel z definovaných údajov bol následne vztiahnutý k veľkosti populácie stanovenej v predošlom kroku, opäť vložený do vhodného rozsahu hodnôt veľkosti populácie a následne zapísaný do databázy.
Stanovenie výmery biotopov v územiach Nátura 2000 bolo vykonané na základe databázy SDF, z ktorej boli v prvom kroku územia rozdelené na alpský a panónsky bioregión. Následne podľa prerátania výmery z percent pokryvnosti daného biotopu v UEV boli zosumarizované podľa jednotlivých biotopov za bioregión spolu a po sčítaní vložené do databázy. Následne boli zadefinované vhodné rozsahy jednotlivých údajov.
Pri vytváraní nových GIS vrstiev bol použitý prístup transformácie pôvodného reportingu, doplneného o nové dáta, ktoré pribudli do databázy za obdobie posledných 6 rokov. Transformácia prebiehala na rôznych úrovniach podľa predošlých podkladov. V prípade biotopov a rastlín išlo o pôvodné vrstvy v sieti 1 x 1 km, v prípade živočíchov bol výskyt v sieti databanky fauny Sloevneska (DFS). Nový formát určuje štandard a požaduje všetky vrstvy rozšírenia v sieti ETRS 10 x 10 km. Z uvedeného dôvodu, boli všetky vrstvy pretransformované, avšak pri transformácii jednej siete do druhej vznikajú odchýlky, ktoré nie je možné eliminovať. Po vytvorení nových vrstiev z podkladu pôvodného reportingu bola každá vrstva revidovaná, odstránené evidentné chyby a staré údaje. Ak to bolo možné, následne boli pridávané nové údaje pre každý druh/biotop zvlášť na základe nových poznatkov za posledné reportingové obdobie. Pre každý novo pridaný GRID (štvorec) existuje podkladový údaj, prípadne zdôvodnenie, rozdielu oproti minulému reportovaciemu obdobiu, kedy mnohokrát nebolo možné identifikovať podkladové dáta, alebo existovali len nepresné údaje.
Cieľom úvodnej fázy bola príprava databázy tak, aby všetky záznamy mali vyplnené potrebné povinné polia a pripravené mapy (vrstvy) rozšírenia v požadovanom formáte.
1.2.2 Prostredná fáza prípravy
Po príprave databázy v novom formáte a GIS vrstiev výskytu druhov a biotopov prebiehalo pripomienkovanie expertmi na národnej úrovni. Expertom boli pripravené všetky potrebné podklady a to konkrétne rozdelené databáza s predvyplnenými údajmi, mapy rozšírenia v online prehliadači a taktiež *.jpeg exporty jednotlivých máp. Vytvorené boli podrobné inštrukcie, podľa ktorých experti pripomienkovali databázovú aj mapovú časť. Do aktívneho pripomienkovania bolo prostredníctvom projektu OPŽP „Príprava a zavedenie monitoringu druhov a biotopov a zlepšenie informovania verejnosti (ďalej len „ŠF Monitoring") zapojených viac ako 30 národných expertov, ktorí boli prostredníctvom zmluvného dodávateľa oslovení a následne kontrahovaní na pripomienkovanie špecificky pridelených biotopov/druhov podľa oblasti ich špecializácie. Experti vpisovali svoje pripomienky priamo do databázy a taktiež priamo tam zdôvodňovali svoje návrhy na jednotlivé zmeny.
Pripomienky k mapám rozšírenia vkladali do pripravených *.xls súborov pre jednotlivé druhy a biotopy, do ktorých vpisovali čísla GRIDov na pridanie, odstránenie a zdôvodnenie navrhovaných zmien. V prípade lesných biotopov ich distribúciu navrhlo Národné lesnícke centrum (NLC) na základe lesníckej typológie a podľa požiadavky ŠOP SR aj na základe kritérií nového metodického pokynu „Mapovanie lesných biotopov" (ŠOP SR, jún 2013). Výmery lesných biotopov boli predmetom osobitného rokovania ŠOP SR s NLC. Rovnako, tak ako v prípade expertov pripomienkujúcich cez uvedený projekt, mali možnosť pripomienkovať aj členovia novo zriadenej koordinačnej rady pre reporting podľa čl. 17 smernice o biotopoch. Z koordinačnej rady sa pripomienkovania zúčastnili len experti, ktorých zastrešoval aj uvedený projekt. Ostatní členovia koordinačnej rady sa k podkladom nevyjadrili. Ďalej boli pripravené podklady samostatne pre veľké šelmy, ktoré boli zaslané na pripomienkovanie rade pre veľké šelmy. Obdržali sme pripomienky od troch členov. Po obdržaní pripomienok od jednotlivých externých expertov, boli postupne zmeny zapracovávané do databázy, aj do máp rozšírenia. V rámci databázy došlo k viac ako 6000 zmenám (z toho cca 3000 vecných), v rámci pripomienkovania máp rozšírenia sa jednalo o viac ako 1000 úprav.
Po zapracovaní pripomienok od externých expertov, boli spracované podklady zaslané na pripomienkovanie jednotlivým organizačným útvarom ŠOP SR. Pripomienky zaslalo viac než 20 expertov ŠOP SR, ktorých pripomienky boli zohľadnené. Vyjadrovali sa predovšetkým k mapám rozšírenia.
Z uvedeného vyplýva, že do prípravy terajšieho reportu bolo zapojených viac ako 50 expertov na národnej úrovni.
1.2.3 Finálna fáza prípravy
Vo finálnej fáze prípravy boli vykonané predovšetkým porovnania zmien, ktoré nastali oproti predošlému obdobiu vo väčšine parametrov. Po identifikovaní zmien boli vykonané potrebné zásahy do databázy kvôli korektnosti údajov. V nadväznosti na to boli vykonané aj logické kontroly, ktoré pomohli identifikovať ešte neupravené nezrovnalosti.
Vykonané logické kontroly:
Druhy
1. Veľkosť populácie nesmie byť väčšia ako veľkosť populácie v územiach Nátura 2000.
2. Kontrola pokryvnosti populácií druhov v územiach Nátura 2000.
3. Priaznivá referenčná populácia nesmie byť väčšia ako skutočná populácia, ak je celkové hodnotenie stavu populácie v priaznivom stave.
4. Priaznivý referenčný areál nesmie byť väčší ako areál, ak je celkové hodnotenie stavu areálu v priaznivom stave.
5. Biotop druhu nesmie byť väčší ako jeho areál.
6. Kontrola celkového hodnotenia stavov podľa hodnotiaceho matrixu (pravidiel definovaných v metodike EK).
7. Kvalita údajov vysoká a zároveň klesajúci trend.
8. Mapa areálu zahŕňa všetky GRIDy mapy rozšírenia.
Biotopy:
1. Veľkosť pokryvnosti biotopu nesmie byť väčšia ako jeho areál.
2. Pokryvnosť v územiach Nátura 2000 nesmie byť väčšia ako pokryvnosť v bioregióne.
3. Kontrola pokryvnosti biotopov v územiach Nátura 2000
4. Priaznivý referenčný areál nesmie byť väčší ako skutočný areál, ak je celkové hodnotenie stavu areálu v priaznivom stave.
5. Priaznivá referenčná pokryvnosť nesmie byť väčšia ako pokryvnosť v bioregióne, ak je hodnotenie pokryvnosti je v priaznivom stave.
6. Kontrola matrixu a celkového hodnotenia stavov, či sú v súlade s pravidlami.
7. Kvalita údajov vysoká a zároveň klesajúci trend.
8. Mapa areálu zahŕňa všetky GRIDy mapy rozšírenia.
Po vykonaní logických kontrol boli vykonané ďalšie úpravy, tak aby bola logika jednotlivých údajov v databáze správna.
V rámci finálnej fázy boli pripravené taktiež mapy areálu. Pri ich tvorbe bola základom už finálna mapa rozšírenia, z ktorej boli prostredníctvom GIS nástroja „range tool" vygenerované mapy areálu. Pri každom druhu/biotope boli zvažované jeho ekologické faktory a podľa toho mu bol prisúdený vhodný areálový variant. Pre vyplnenie medzier (Gap) boli použité hodnoty 0, 1, 2 alebo 3, podľa špecifík jednotlivého druhu alebo biotopu. V prípade vyšších rastlín „range tool" použitý nebol, vzhľadom na pomerne dobré údaje o areáli, ktoré by sa prostredníctvom tohto nástroja iba znepresnili.
2 Všeobecné informácie
2.1 Počet území európskeho významu (SCI a SAC)
Nižšie uvedená tabuľka uvádza celkový počet a celkovú rozlohu území navrhnutých a vyhlásených podľa smernice o biotopoch (lokality významné pre Spoločenstvo - SCI a osobitných oblastí ochrany - SAC), súhrnnú rozlohu lokalít.
|
|
Spolu |
|
|
Počet |
Rozloha (Ha) |
|
|
SCIs & SACs |
473 |
584349.84 |
|
SACs * |
305 |
384933.17 |
*stav k 1.1.2014

2.2 Počet území s komplexnými programami starostlivosti (článok 6 (1))
Počet území, pre ktoré boli prijaté komplexné programy starostlivosti: 8
Percento plochy území, ktorá je pokrytá komplexnými programami starostlivosti: 1%
Počet území, pre ktoré programy starostlivosti sú v príprave: 131
3 Počet biotopov a druhov európskeho významu
Tabuľka v tejto časti uvádza počet typov biotopov a druhov / poddruhov v každej prílohe smernice o biotopoch podľa biogeografických oblastí na Slovensku.
|
Bioregión
|
Biotopy |
Druhy |
||||||
|
Príloha I |
Príloha II |
Príloha IV |
Príloha V |
|||||
|
Neprioritné |
Prioritné |
Neprioritné |
Prioritné |
Vrátane druhov v prílohe II |
Bez druhov v prílohe II |
Vrátane druhov v prílohe II |
Bez druhov v prílohe II |
|
|
Počet biotopov a druhov |
44 |
22 |
110 |
23 |
138 |
44 |
24 |
15 |
|
67 |
133 |
138 |
24 |
|||||
|
Alpský |
41 |
16 |
92 |
21 |
117 |
39 |
21 |
14 |
|
Panónsky |
28 |
16 |
89 |
8 |
109 |
40 |
21 |
13 |
Počet okrajových a sporadických výskytov biotopov: 0
Počet okrajových a sporadických výskytov druhov: 4
Počet druhov vyhynutých pred prijatím smernice o biotopoch: 23
Počet druhov vyhynutých po prijatí smernice o biotopoch: 0
4 Informácia o stave biotopov a druhov európskeho významu
4.1 a) Celkové hodnotenie stavu biotopov a druhov eur. významu (%)
Nasledovné grafy zobrazujú stav biotopov a druhov eur. významu v Slovenskej republike v kategóriách priaznivého stavu (FV, U1, U2 a XX) a porovnanie stavu s predošlou periódou reportingu. Informácie, na ktorých sú založené údaje na obrázkoch sú uvedené v tabuľke.
|
||||||
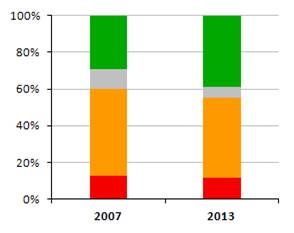
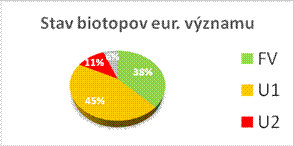
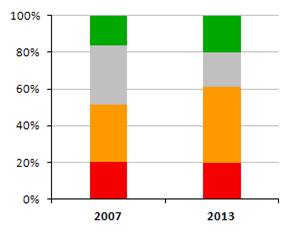
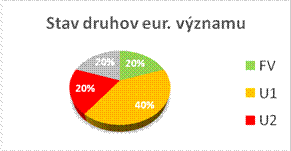
|
|||||||||||||||||||||||||||||||||||
4.1 b) Percento hodnotení zmien stavu medzi jednotlivými periódami podávania správy
Nasledujúca tabuľka ukazuje percentuálne hodnotenie zmien v stave biotopov a druhov medzi dvoma reportingovými periódami (2001-2006 a 2007-2013) a percentá všetkých zmien, kde zmena bola označená ako skutočná zmena stavu v realite a nie len na základe lepších poznatkov. Údaje boli prevzaté z tzv "audit trail ", kde Slovenská republika zdôvodnila podstatu zmien. Podrobné výsledky z tzv. “audit trail” sú uvedené v kapitole 8.
|
4.2 Zlepšujúci sa/zhrošujúci sa trend stavu biotopov a druhov eur. významu v nepriaznivom stave. (%)
Nasledovné grafy uvádzajú pomer hodnotení v nepriaznivom stave (U1 & U2), ktoré sa zlepšujú, zhoršujú, sú stabilné alebo ich trend stavu nie je známy.
|
||||
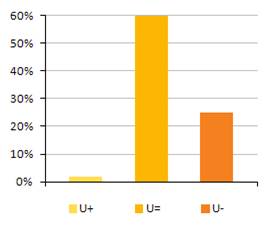
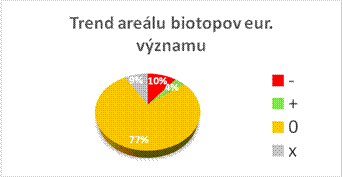
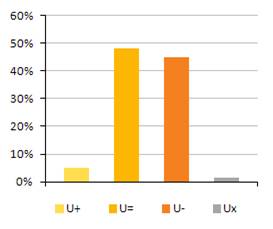
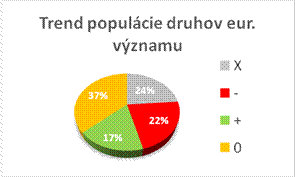
Nasledovná tabuľka uvádza trendy v stave biotopov a druhov eur. významu samostatne v tých prípadoch, kde bolo hodnotenie stavu nepriaznivé – nevyhovujúce (U1) a nepriaznivé – zlé (U2).
|
Poznámka: U1+ = nepriaznivý – nevyhovujúci, ale zlepšujúci sa, U1= = nepriaznivý – nevyhovujúci, ale stabilný, U1- = nepriaznivý – nevyhovujúci a zhoršujúci sa, U1x = nepriaznivý - nevyhovujúci a trend stavu je neznámy, U2+ = nepriaznivý – zlý, ale zlepšujúci sa, U2= = nepriaznivý – zlý, ale stabilný, U2- = nepriaznivý – zlý a zhoršujúci sa, U2x = nepriaznivý - zlý a trend stavu je neznámy
4.3 Celkové hodnotenie stavu biotopov a druhov eur. významu podľa jednotlivých biogeografických regiónov (%)
Nasledovné grafy uvádzajú percentuálne hodnotenie stavu podľa biogeografických regiónov samostatne pre biotopy a druhy eur. významu.
|
||
|
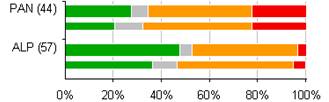
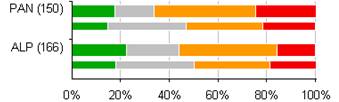
Poznámka: širšia línia v grafe odpovedá perióde reportingu za roky 2007 – 2012, užšia línia pod ňou zobrazuje údaje z reportingovej periódy za roky 2004 – 2006. Počet v zátvorkách uvádza počet hodnotení stavu v jednotlivých bioregiónoch v perióde rokov 2007 – 2012.

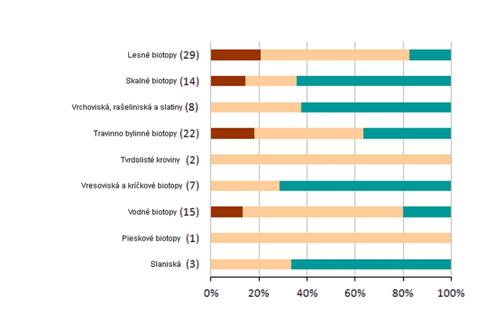
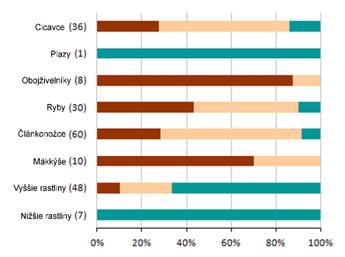
4.4 Celkové hodnotenie stavu podľa jednotlivých skupín biotopov/druhov (%)
Nasledovné grafy ukazujú percento hodnotenia podľa skupín biotopov a druhov. Na hornej širšej lište sú zobrazené údaje za reportingovú periódu 2007 – 2012 a za roky 2004- 2006 na spodnej užšej lište. Sumárne číselné informácie na základe ktorých je graf vytvorený sú uvedené v tabuľke pod grafom.
|
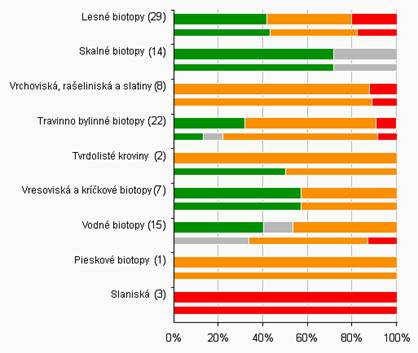
Poznámka: širšia línia v grafe odpovedá perióde reportingu za roky 2007 – 2012, užšia línia pod ňou zobrazuje údaje z reportingovej periódy za roky 2004 – 2006. Počet v zátvorkách uvádza počet hodnotení stavu v jednotlivých bioregiónoch v perióde rokov 2007 – 2012.
Slaniskové biotopy patria medzi najohrozenejšie biotopy v celoeurópskom meradle. Ich stav je na Slovensku hodnotený ako nepriaznivý-zlý. Rovnako aj čiastkové hodnotenia areálu, štruktúry a funkcií, i vyhliadky do budúcnosti sú hodnotené ako nevyhovujúce alebo zlé.
Vrchoviská, rašeliniská a slatiny sú biotopy, u ktorých zmeny vo vodnom režime a nedostatočný manažment spôsobujú zhoršovanie stavu až degradáciu biotopov. Ich stav je na Slovensku hodnotený ako nepriaznivý-nevyhovujúci až nepriaznivý-zlý.
Ďalšou skupinou biotopov, ktorej stav je hodnotený ako nepriaznivý-nevyhovujúci, sú pieskové biotopy vyskytujúce sa u nás len na Borskej nížine na nespevnených kyslých pieskových dunách. Ohrozené sú zalesňovaním a sukcesiou.
Tvrdolisté kroviny sú reprezentované biotopom porastov s borievkou obyčajnou. Ich stav sa v porovnaní s predchádzajúcim reportovaným obdobím zmenil na nepriaznivý-nevyhovujúci. Súvisí to s nepriaznivo hodnotenými vyhliadkami do budúcnosti, ktoré vychádzajú z vplyvov sukcesie a čistením extenzívne využívaných trávnych porastov od náletových drevín a krov.
Vodné biotopy (horské vodné toky, stojaté vody) závislé od dynamiky vodných tokov a primeranej kvality, vresoviská a kríčkové biotopy závislé od občasného a cieleného manažmentu a travinno-bylinné biotopy vyžadujúce pravidelný manažment sa nachádzajú medzi hodnotením stavu priaznivý až nepriaznivý-zlý.
Relatívne najmenej ohrozenou skupinou sú skalné a sutinové biotopy a jaskyne.

|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
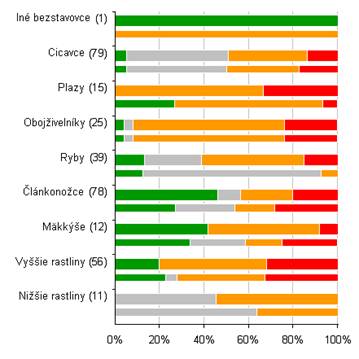
Poznámka: širšia línia v grafe odpovedá perióde reportingu za roky 2007 – 2012, užšia línia pod ňou zobrazuje údaje z reportingovej periódy za roky 2004 – 2006. Počet v zátvorkách uvádza počet hodnotení stavu v jednotlivých bioregiónoch v perióde rokov 2007 – 2012.

|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
4.4.2.1 Nižšie a vyššie rastliny
 Lepší stav
zachovania dosahujú rastliny, ktoré sa vyskytujú v alpskom bioregióne (stredná
a severná časť Slovenska), čo vyplýva z menšieho narušenia prírodného
prostredia. K najviac ohrozeným druhom aj naďalej patria napr. jesienka
piesočná (Colchicum arenarium) a kosatec piesočný (Iris humilis
subsp. arenaria), ktoré na Slovensku rastú len na jedinej lokalite (s
viacerými mikrolokalitami) v blízkosti Štúrova, na severnej hranici svojho
areálu. Jedinú lokalitu na Slovensku majú aj kosienka karbincolistá (Serratula
lycopifolia) či mečík močiarny (Gladiolus palustris). Väčšina
druhov, ktoré sa nachádzajú v nepriaznivom stave ochrany, sa vyskytuje na veľmi
špecifických a zraniteľných typoch biotopoch (rašeliniská, slatiny, podmáčané
lúky, viate piesky, atď.). Najväčšiu skupinu tvoria rastliny, ktoré sa
nachádzajú v nevyhovujúcom stave. V priaznivom stave ochrany sa na Slovensku
nachádzajú druhy ako snežienka jarná (Galanthus nivalis), zvonček
hrubokoreňový (Campanula serrata) a ďalšie, ktoré sú v západnej Európe
už pomerne vzácne alebo sa tam už vôbec nevyskytujú, napr. cyklámen fatranský (Cyclamen
fatrense), lyžičník tatranský (Cochlearia tatrae), či vrchovka
alpínska (Tozzia carpathica). V rozpore s pomerne jasným hodnotením vyšších
rastlín je v súčasnosti pri 45 % neznámy stav zachovania machorastov (nižších
rastlín). Zavedený systematický
monitoring zlepšuje poznanie nižších rastlín eur. významu vo významnej miere
a predpokladáme, že v ďalšom reportingovom kole už budú z hľadiska
stavu hodnotené všetky druhy nižších rastlín eur. významu.
Lepší stav
zachovania dosahujú rastliny, ktoré sa vyskytujú v alpskom bioregióne (stredná
a severná časť Slovenska), čo vyplýva z menšieho narušenia prírodného
prostredia. K najviac ohrozeným druhom aj naďalej patria napr. jesienka
piesočná (Colchicum arenarium) a kosatec piesočný (Iris humilis
subsp. arenaria), ktoré na Slovensku rastú len na jedinej lokalite (s
viacerými mikrolokalitami) v blízkosti Štúrova, na severnej hranici svojho
areálu. Jedinú lokalitu na Slovensku majú aj kosienka karbincolistá (Serratula
lycopifolia) či mečík močiarny (Gladiolus palustris). Väčšina
druhov, ktoré sa nachádzajú v nepriaznivom stave ochrany, sa vyskytuje na veľmi
špecifických a zraniteľných typoch biotopoch (rašeliniská, slatiny, podmáčané
lúky, viate piesky, atď.). Najväčšiu skupinu tvoria rastliny, ktoré sa
nachádzajú v nevyhovujúcom stave. V priaznivom stave ochrany sa na Slovensku
nachádzajú druhy ako snežienka jarná (Galanthus nivalis), zvonček
hrubokoreňový (Campanula serrata) a ďalšie, ktoré sú v západnej Európe
už pomerne vzácne alebo sa tam už vôbec nevyskytujú, napr. cyklámen fatranský (Cyclamen
fatrense), lyžičník tatranský (Cochlearia tatrae), či vrchovka
alpínska (Tozzia carpathica). V rozpore s pomerne jasným hodnotením vyšších
rastlín je v súčasnosti pri 45 % neznámy stav zachovania machorastov (nižších
rastlín). Zavedený systematický
monitoring zlepšuje poznanie nižších rastlín eur. významu vo významnej miere
a predpokladáme, že v ďalšom reportingovom kole už budú z hľadiska
stavu hodnotené všetky druhy nižších rastlín eur. významu.
 4.4.2.2
Bezstavovce
4.4.2.2
Bezstavovce
4.4.2.2.1 Motýle
Zo skupiny motýľov najhoršie hodnotenie stavu má druh žltáčik zanoväťový (Colias myrmidone) (U2- ALP/PAN), druh očkáň mätonohý (Lopinga achine) (U2 ALP/PAN) a druh modráčik čiernoškvrnný (Maculinea arion) (U2- PAN). Naopak v priaznivom stave boli vyhodnotené napr. druhy ako napr. spriadač kostihojový (Callimorpha quadripunctaria) (FV ALP/PAN), priadkovec trnkový (Eriogaster catax) (FV ALP/PAN), modráčik bahniskový (Maculinea nausithous) (FV ALP/PAN), jasoň chochlačkový (Parnassius mnemosyne) (FV ALP/PAN) a niektoré ďalšie druhy motýľov eur. významu. Hodnotenie stavu do určitej miery odráža aj odhadovaná veľkosť populácie uvedená v nasledovnom grafe. Dôležité z hľadiska ochrany druhov sú určite tie druhy, ktoré majú odhadovanú početnosť pod 15000 jedincov.

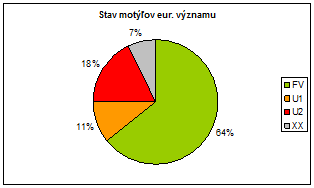
Celkový stav motýľov eur. významu je na Slovensku podľa hodnotenia v správe pomerne pozitívny, pretože väčšina hodnotení druhov je v stave priaznivom. (64 %). Avšak je potrebné podotknúť, že 18 % hodnotení je v stave nepriaznivom – zlom (U2), ktoré jednoznačne indikujú potrebu zlepšenia stavu predovšetkým týchto druhov.
4.4.2.2.2 Chrobáky
 U chrobákov sú v zlom stave niektoré druhy viazané na mŕtve drevo ako
napr. boros Schneiderov (Boros schneideri) (U2- ALP), kováčik
fialový (Liminiscus violaceus) (U2- PAN), drevník ryhovaný (Rhysodes
sulcatus) (U2- PAN) a ďalšie. V priaznivom stave sa nachádzajú napr.
druhy plocháč červený (Cucujus cinnaberinus), behúnik maďarský (Duvalius
hungaricus), roháč veľký (Lucanus cervus). Z hľadiska veľkosti
populácie odhady medzi jednotlivými druhmi sú pomerne odlišné, od malých
populácií druhu borosa Schneidrovho (Boros schneideri), bystrušky
Zawadského (Carabus zawadzkii) až po väčšie populácie menej vzácnych,
ale stále významných druhov pre ekosystémy ako napr. roháča veľkého (Lucanus
cervus) a ďalších druhov. Z hľadiska priorít pre ochranu je potrebné
sledovať populácie, ktoré sú odhadované na 10000 jedincov a menej.
U chrobákov sú v zlom stave niektoré druhy viazané na mŕtve drevo ako
napr. boros Schneiderov (Boros schneideri) (U2- ALP), kováčik
fialový (Liminiscus violaceus) (U2- PAN), drevník ryhovaný (Rhysodes
sulcatus) (U2- PAN) a ďalšie. V priaznivom stave sa nachádzajú napr.
druhy plocháč červený (Cucujus cinnaberinus), behúnik maďarský (Duvalius
hungaricus), roháč veľký (Lucanus cervus). Z hľadiska veľkosti
populácie odhady medzi jednotlivými druhmi sú pomerne odlišné, od malých
populácií druhu borosa Schneidrovho (Boros schneideri), bystrušky
Zawadského (Carabus zawadzkii) až po väčšie populácie menej vzácnych,
ale stále významných druhov pre ekosystémy ako napr. roháča veľkého (Lucanus
cervus) a ďalších druhov. Z hľadiska priorít pre ochranu je potrebné
sledovať populácie, ktoré sú odhadované na 10000 jedincov a menej.
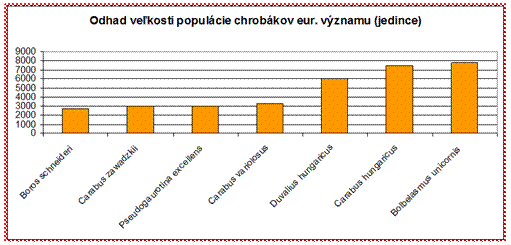
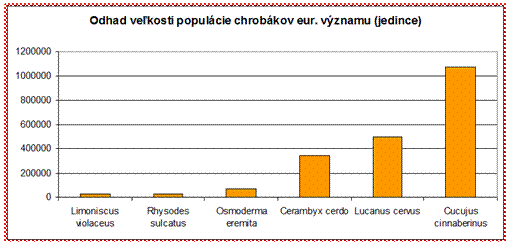
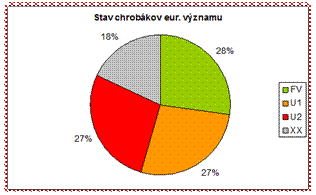
Z hľadiska hodnotenia stavu je väčšina druhov chrobákov eur. významu v nepriaznivom stave (54 %), čo je pomerne negatívna situácia a bude potrebné významne podporiť druhy aktívnym manažmentom a lepším zabezpečením ochrany pre túto ohrozenú skupinu.

4.4.2.2.3 Vážky
V prípade vážok bolo hodnotených len 5 druhov. V zlom stave boli hodnotené druhy vážka jednoškvrnná (Leucorrhinia pectoralis) (U2 ALP) a šidielko ozdobné (Coenagrion ornatum) (U2 PAN). V priaznivom stave boli vyhodnotené druhy pásikavec veľký (Cordulegaster heros) (FV ALP), vážka jednoškvrnná (Leucorrhinia pectoralis) (FV PAN) a klinovka žltonohá (Stylurus flavipes) (FV PAN). Z hľadiska odhadu veľkosti populácie početnejšie populácie majú druhy klinovka hadia (Ophiogomphus cecilia) a klinovka žltonohá (Stylurus flavipes). Naproti tomu druhy šidielko ozdobné (Coenagrion ornatum), pásikavec veľký (Cordulegaster heros) a vážka jednoškvrnná (Leucorhinia pectoralis) majú odhadovanú populáciu menšiu ako 10000 jedincov.
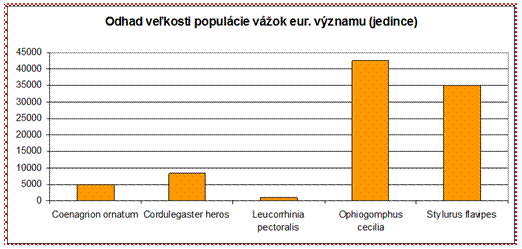
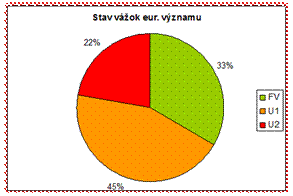
Pomer priaznivého/nepriaznivého stavu v prípade vážok eur. významu je v ich neprospech a teda väčšina hodnotení je uvádzaných v správe v stave nepriaznivom. Z tohto ohľadu je potrebné zabezpečiť zlepšenie stavu do budúcnosti a zvrátiť tak aktuálne negatívne hodnotenie.
4.4.2.2.4 Rovnokrídlovce
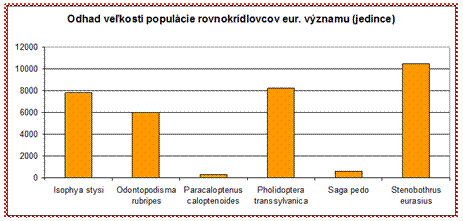
Z rovnokrídlovcov eur. významu sú v priaznivom stave hodnotené napr. druhy kobylka Štysova (Isophya stysi) (FV ALP/PAN), koník východný (Odontopodisma rubripes) (FV ALP/PAN). Pozitívne je, že žiadny z druhov nebol vyhodnotený v zlom stave (U2). V nevyhovujúcom stave bol vyhodnotený napr. druh sága stepná (Saga pedo) (U1 ALP/PAN). Z hľadiska odhadu veľkosti populácie nízkopočetné populácie sú odhadované pri druhoch sága stepná (Saga pedo) a koník Brunnerov (Paracaloptenus caloptenoides), avšak ani u druhov koník slovanský (Stenobothrus eurasius) a kobylka sedmohradská (Pholidoptera transsylvanica) odhadovaný počet jedincov nepresahuje veľkosť 12 000 jedincov a preto je potrebné im venovať náležitú pozornosť.
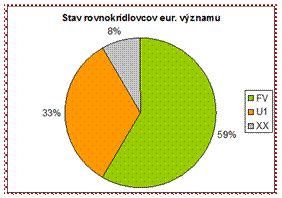
Pomer priaznivého stavu ku nepriaznivému je v prípade rovnokrídlovcov pozitívny a teda pravládajú hodnotenia v stave priaznivom. (59 %)
 4.4.2.2.5 Mäkkýše
4.4.2.2.5 Mäkkýše
V priaznivom stave v alpskom aj panónskom bioregióne boli hodnotené dva druhy – pimprlík mokraďný (Vertigo angustior) a slimák záhradný (Helix pomatia). Uvedené druhy majú zároveň aj najväčšiu odhadovanú početnosť populácie. V zlom stave v panónskom bioregióne bol vyhodnotený druh pimprlík bruškatý (Vertigo moulinsiana), ktorý spolu s druhom pimprlík močiarny (Vertigo geyeri) majú zároveň najmenšiu odhadovanú veľkosť populácie.
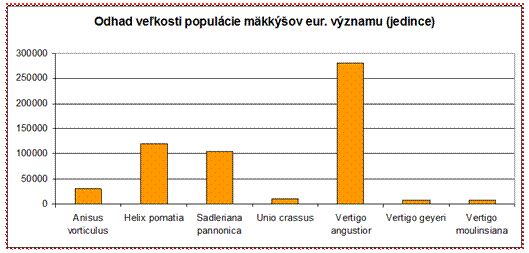
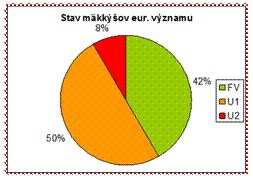
Z hľadiska celkového vyhodnotenia stavu však prevládajú negatívne hodnotenia a teda väčšina druhov má hodnotenia v stave nepriaznivom. (58 %)
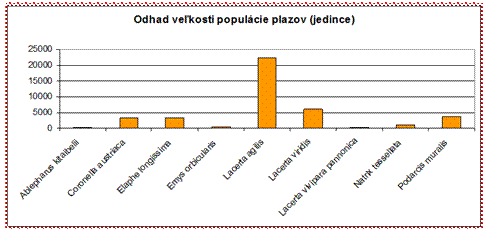
4.4.2.3 Plazy a obojživelníky
 Celkový stav zachovania plazov a obojživelníkov
vyplývajúci z údajov z reportingu je veľmi negatívny. Žiadny z hodnotených
druhov plazov eur. významu nebol vyhodnotený v stave priaznivom.
Celkový stav zachovania plazov a obojživelníkov
vyplývajúci z údajov z reportingu je veľmi negatívny. Žiadny z hodnotených
druhov plazov eur. významu nebol vyhodnotený v stave priaznivom.
Až 3 druhy plazov sú v zlom stave (U2). Všetky ostatné plazy sú hodnotené v stave nevyhovujúcom (U1). Sú to druhy ako napr. jašterica múrova (Podarcis muralis), ktorá je ohrozená sukcesiou a upustením od pôvodných foriem hospodárenia rovnako ako jašterica zelená (Lacerta viridis), kde zarastaním xerotermných krovín dochádza k úbytku vhodných biotopov. V panónskom bioregióne je v nepriaznivom stave (U2) aj korytnačka močiarna (Emys orbicularis).
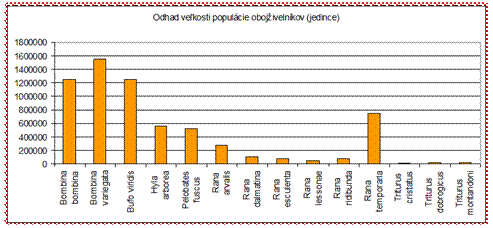
 Pri obojživelníkoch
je situácia podobná. Skokan hnedý (Rana temporaria) je v priaznivom
stave len v alpskom bioregióne, v panónskom mu bol prisúdený stav nevyhovujúci.
Všetky ostatné druhy sú v nevyhovujúcom alebo zlom stave! Nepriaznivé závery
stavu biotopov, v ktorých plazy a obojživelníky žijú, sú takmer totožné s
celkovými závermi stavu druhov, čo naznačuje signifikantnú viazanosť plazov a
obojživelníkov na svoje prostredie a jeho kvalitu.
Pri obojživelníkoch
je situácia podobná. Skokan hnedý (Rana temporaria) je v priaznivom
stave len v alpskom bioregióne, v panónskom mu bol prisúdený stav nevyhovujúci.
Všetky ostatné druhy sú v nevyhovujúcom alebo zlom stave! Nepriaznivé závery
stavu biotopov, v ktorých plazy a obojživelníky žijú, sú takmer totožné s
celkovými závermi stavu druhov, čo naznačuje signifikantnú viazanosť plazov a
obojživelníkov na svoje prostredie a jeho kvalitu.
Výsledky slovenského reportingu vôbec nie sú prekvapujúce. Aj v rámci celej Európy sa javí skupina plazov a obojživelníkov ako najviac ohrozená spomedzi všetkých druhov európskeho významu a je potrebné venovať im nemalú pozornosť.
Čiastkové závery teda naznačujú, že vzhľadom na silnú viazanosť týchto druhov na biotop, by mali byť manažmentové opatrenia zamerané práve na zlepšenie kvantity a kvality biotopov, v ktorých sa plazy a obojživelníky vyskytujú.
4.4.2.4 Ryby
Zo skupiny rýb sa v priaznivom stave nachádza len boleň dravý (Aspius aspius), lopatka dúhová (Rhodeus sericeus amarus) a hlaváč bieloplutvý (Cottus gobio) v alpskom bioregióne, ktorých populácie a areál boli hodnotené ako vyhovujúce. Referenčná (cieľová) veľkosť populácie a areál všetkých ostatných druhov rýb však majú byť väčšie ako v súčasnosti. Celkový stav je klasifikovaný pri mnohých druhoch ako neznámy.
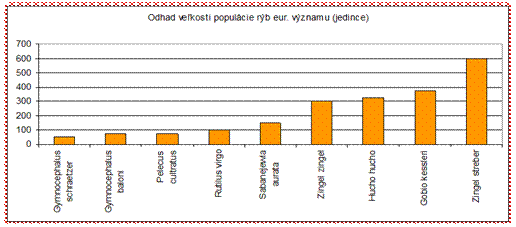
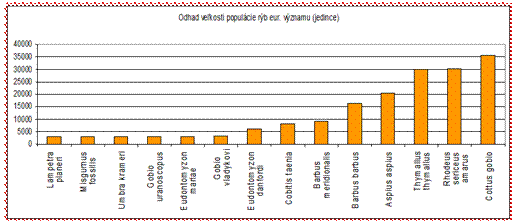
Alarmujúca je situácia s niektorými v minulosti bežnými druhmi rýb (hlavátka, lipeň, mrena), ktorých populácie sú v súčasnosti ohrozované najmä reguláciami tokov, výstavbou bariér (malé vodné elektrárne) a predáciou kormorána veľkého. Z kriticky ohrozených druhov sa na Slovensku zatiaľ vyskytujú mihule (Lampetra planeri, Eudontomyzon danfordi, E. mariae), kolky (Zingel streber, Z. zingel), hrúz Kesslerov (Gobio kesslerii), hrúz fúzatý (Gobio uranoscopus), blatniak tmavý (Umbra krameri) a čík európsky (Misgurnus fossilis), ktorých výskyt sa viaže len na málo slovenských tokov.
Napriek tomu, že sa v poslednom desaťročí v našich tokoch zlepšila čistota vody, v oblasti revitalizácie poškodených úsekov tokov a spriechodnenia riek máme obrovské rezervy.
Všeobecne môžeme povedať, že chránené druhy rýb sú malo preskúmané, preto bude nevyhnutné venovať im väčšiu pozornosť.
4.4.2.5 Cicavce
Hodnotených bolo 48 druhov cicavcov. S výnimkou všetkých u nás zaznamenaných 28 druhov netopierov a niektorých druhov hlodavcov, sa jedná o najlepšie preskúmanú skupinu živočíchov.
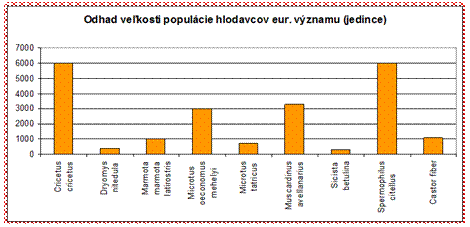
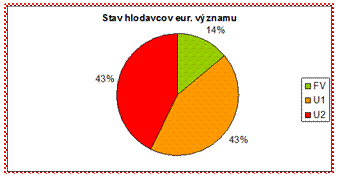
4.4.2.5.1 Hlodavce
V rámci skupiny hlodavcov je v priaznivom stave vyhodnotený len druh bobor vodný (Castor fiber). Všetky ostatné druhy hlodavcov sú v nepriaznivom stave, čo je pomerne negatívny výsledok.

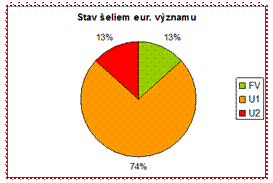
4.4.2.5.2 Šelmy

Zo šeliem eur. významu boli v priaznivom stave hodnotené len druhy medveď hnedý (Ursus arctos) a vlk dravý (Canis lupus) v alpskom bioregióne. Všetky ostatné druhy boli vyhodnotené v nepriaznivom stave. V zlom stave (U2) bol vyhodnotený druh tchor stepný (Mustela eversmanii).
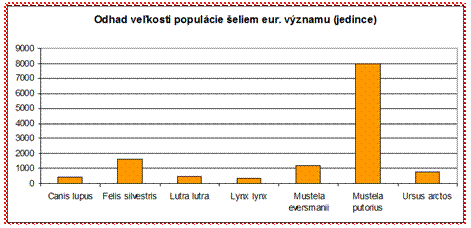

*Martes martes – 67500 jedincov
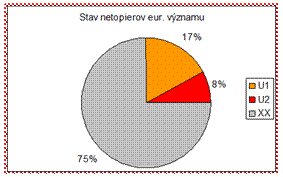
4.4.2.5.3 Netopiere
Stav väčšiny druhov netopierov bol hodnotený ako neznámy. Napriek realizácii čiastkového monitoringu netopierov je odhad ich populácií veľmi problematický a predpokladáme, že v nasledujúcom období už stav netopierov bude vyhodnotený. V zlom stave (U2) boli vyhodnotené druhy netopier sťahovavý (Miniopterus schreibersii) a netopier hryzavý (Nyctalus noctula).

Odhad početnosti netopierov eur. významu (jedince)
|
Myotis alcathoe |
1 |
|
Hypsugo savii |
30 |
|
Nyctalus lasiopterus |
50 |
|
Pipistrellus kuhlii |
125 |
|
Pipistrellus pygmaeus |
150 |
|
Pipistrellus nathusii |
325 |
|
Vespertilio murinus |
375 |
|
Nyctalus leisleri |
600 |
|
Myotis bechsteinii |
1050 |
|
Myotis nattereri |
1050 |
|
Rhinolophus ferrumequinum |
2000 |
|
Eptesicus nilssonii |
3000 |
|
Myotis brandtii |
3025 |
|
Myotis blythii |
3300 |
|
Myotis mystacinus |
3300 |
|
Plecotus auritus |
3300 |
|
Myotis daubentonii |
6000 |
|
Nyctalus noctula |
6000 |
|
Plecotus austriacus |
6000 |
|
Myotis emarginatus |
7250 |
|
Rhinolophus euryale |
7500 |
|
Myotis dasycneme |
7525 |
|
Miniopterus schreibersii |
8000 |
|
Rhinolophus hipposideros |
10750 |
|
Eptesicus serotinus |
30000 |
|
Barbastella barbastellus |
78000 |
|
Myotis myotis |
157500 |
|
Pipistrellus pipistrellus |
303000 |
4.4.2.5.4 Kopytníky
V rámci skupiny kopytníkov sa hodnotili dva druhy a to kamzík vrchovský tatranský (Rupicapra rupicapra tatrica) a zubor európsky (Bison bonasus). Pozitívnym výsledkom je, že v prípade kamzíka sa jeho stav zlepšil z kategórie nepriaznivý – zlý (U2) na kategóriu nepriaznivý – nevyhovujúci (U1) aj vďaka faktu, že sa za obdobie šiestich rokov od posledného hodnotenia veľkosť populácie takmer zdvojnásobila. Zubor európsky má stanovené hodnotenie stavu ako nepriaznivý – nevyhovujúci (U1).

4.5 Dôvody zmeny v hodnoteniach stavu medzi reportingovými periódami (%)
Nasledovná tabuľka poskytuje prehľad zdôvodnení zmien v paramatroch areál a výmera biotopu, populácia a biotop druhu medzi jednotlivými reportingovými periódami 2004 – 2006 a 2007 – 2012. Tabuľka udáva percento hodnotení biotopov a druhov pre ktoré bol čiaskotvý dôvod zmeny uvedený. Formát reportingu udáva tri rôzne zdôvodnenia zmeny: skutočná zmena stavu, zlepšenie poznatkov a použitie inej metódy pri príprave správy.
|
|||||||||||||||||||||||||||||
Poznámka:. Aj viac ako jeden dôvod môže byť uvedený pre každý biotop alebo druh.
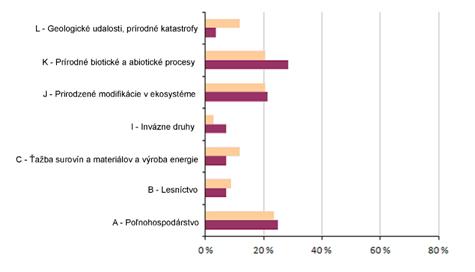
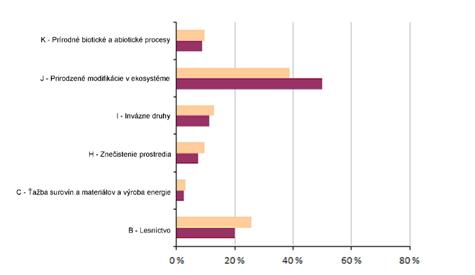
5 Hodnotenie vplyvov a ohrození (%)
Nasledujúca sekcia poskytuje informácie o dôležitosti vplyvov a ohrození (agregovaných do hlavných oblastí) pre reportované biotopy a druhy eur. významu. Grafy znázorňujú percento hodnotení s uvedením aspoň jedného vplyvu/ohrozenia, ktorého dôležitosť bola vyhodnotená ako “vysoká”. Informácie o počte vplyvov a ohrození uvedeného typu sú uvedené v tabuľke pod grafmi.
|
||||||||||||||||||||||||||
|
||||||||||||||||||||||||||
|
|||||||||||||||||||||||
|
|||||||||||||||||||||||

6 Pokryvnosť území Natura 2000 a ochranárske opatrenia
Poznámka: Grafy v sekcii 6 pojednávajú len o biotopoch z prílohy 1 a druhoch z prílohy 2 smernice o biotopoch
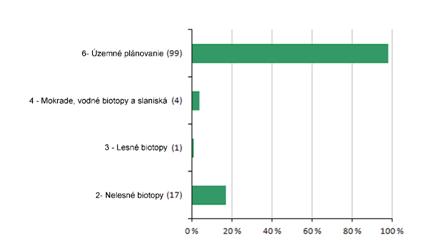
6.1 Pokryvnosť území Natura 2000 (%)
Nasledujúca časť prezentuje štatistiku pokrytia biotopov z prílohy 1 a druhov z prílohy 2 smernice o biotopoch v územiach Natura 2000 podľa jednotlivých skupín biotopov/druhov. Aritmetický priemer bol použitý pri rôznych hodnotách uvedených v poliach minimum/maximum. Informácie o sumárnych hodnotách hodnotení pre pokrytie sieťou Natura 2000 na základe ktorých boli grafy vytvorené sú uvedené v tabuľke pod grafom.
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|||||||||||||||||||||||||||||||||||||||||||||||||
|
|||||||||||||||||||||||||||||||||||||||||||||||||
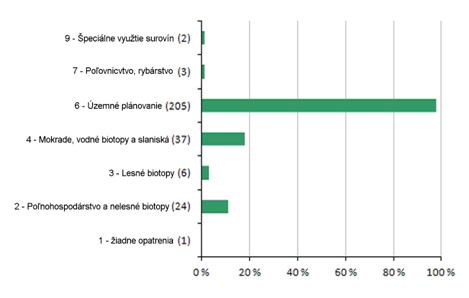
6.2 Hlavné ochranárske opatrenia (%)
Nasledujúca sekcia poskytuje informácie o dôležitosti ochranárskych opatrení implementovaných počas periódy rokov 2007 – 2012 pre biotopy z prílohy 1 a druhy z prílohy 2 smernice o biotopoch. Grafy znázorňujú precento hodnotení, v ktorých bolo uvedené aspoň jedno ochranárske opatrenie s hodnotou “vysoká dôležitosť”. Informácia o sumárnych hodnotách, na základe ktorých boli grafy pripravené sú uvedené v tabuľke pod grafom.
|
|
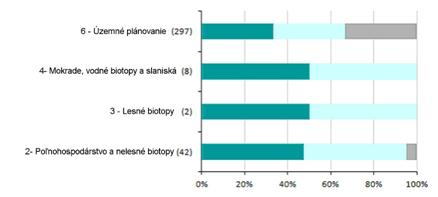
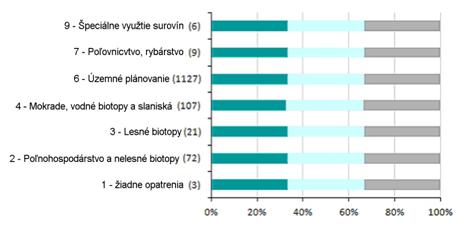
6.3 Vplyv ochranárskych opatrení (%)
Nasledujúca časť poskytuje informácie o efekte implementovaných ochranárskych opatrení. Grafy zobrazujú kategóriu opatrenia a časovú náročnosť ochranárskeho efektu. Súhrnné informácie sú uvedené v tabuľke pod grafom.
|
|
|||||||||||||||||||||||||||||||||||
|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||

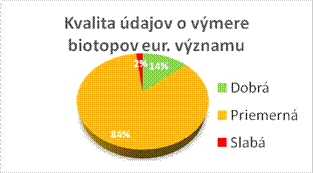
7 Kvalita údajov a kompletnosť
Cieľom tejto sekcie je poskytnúť prehľad o chýbajúcich dátach v reporte, väčšina chýbajúcich údajov je z dôvodu nedostatočných poznatkov. Tabuľky uvádzajú percento hodnotení s chýbajúcimi informáciami pre hodnotenie stavu a závery.
7.1 a) Percento potrebných informácií, ktoré chýbajú v reporte (%)
Biotopy
|
|||||||||||||||||||||||||||||||||||||||
Druhy
|
|||||||||||||||||||||||||||||||||||||||||||||
7.1 b) Percento informácií reportovaných v stave neznámom (%)
Biotopy
|
Druhy
|
7.2 Metódy použité pre stanovenie hodnôt (%)
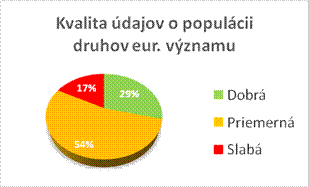
Táto sekcia prezentuje informácie o kvalite údajov, stanovených hodnôt, trendoch v reportoch pre biotopy a druhy. Pre niektoré parametre a trendy formát reportingu vyžaduju indikáciu jednej z troch metód (kompletný prieskum alebo štatisticky robustný odhad, čiastkové údaje s použitím extrapolácie a modelovania, expertný odhad s minimálnimi podkladmi) použitej pri stanovení jednotlivých hodnôt v správe. Tabuľky v tejto sekcii prezentujú percento hodnotení pre biotopy/druhy, pre ktoré boli hodnoty stanovené na základe vyššie spomenutých troch metód.
Biotopy
|
Druhy
|
Zdroje informácií:
Link na všeobecnú časť správy na CDR
Link na report pre biotopy na CDR
Link na report pre druhy na CDR
8. Zoznam reportovaných biotopov a druhov eur. významu a ich stav
V tejto sekcii sú uvedné reportované biotopy a druhy, ich celkový stav v reportingových periódach 2004 – 2006 (uvedená ako rok 2007) a 2007 – 2012 (uvedená ako rok 2013). Tabuľky navyše taktiež obsahujú zdôvodnenie zmeny stavu, v prípade, že sa udiala medzi jednotlivými reportovanými periódami.
Zmeny v porovnaním s predošlou verziou správy za roky 2004 - 2006
Zmeny v stavoch sú najdôležitejšími zmenami v porovnaní s predošlým kolom reportingu. V prípade druhov došlo k zmenám v celkovom hodnotení stavu v 72 prípadoch, z toho v 20 prípadoch ide o zlepšenie stavu, v 11 prípadoch o zhoršenie stavu a v 41 prípadoch o zmenu z neznámeho do priaznivého, nevyhovujúceho alebo zlého stavu.
V prípade biotopov došlo k zmenám v celkovom hodnotení stavu v 16 prípadoch, z toho v 7 prípadoch ide o zlepšenie stavu, v 1 prípade zhoršenie stavu, v 2 prípadoch zmena zo zlého do neznámeho stavu a v 6 prípadoch z neznámeho do priaznivého alebo nevyhovujúceho stavu. Zmeny stavov sú spôsobené najmä zlepšením poznatkov.
Na základe záverov bioseminára konaného v roku 2012 a taktiež na základe nových poznatkov boli viaceré druhy a biotopy navrhnuté na odstránenie z referenčného zoznamu.
Kódy zdôvodnenia zmien uvedených v tabuľke sú nasledovné:
a = skutočná zmena: celkový stav sa zlepšil v dôsledku prírodných alebo neprírodných dôvodov (manažment, intervencie, a pod)
b1 = zmena z dôvodu spresnenia dát (napr. lepšie zmapovanie rozšírenia), alebo zlepšenie znalostí (napr. o ekológii druhu alebo lokalitách)
b2 = zmena z dôvodu taxonomickej revízie
c1 = zmena z dôvodu použitia iných metód pre meranie alebo hodnotenie jednotlivých parametrov alebo celkového stavu
c2 = zmena z dôvodu použitia iných rozsahov pri hodnotení napr. úprava priaznivej referenčnej hodnoty
d = žiadne informácie o povahe zmeny
e = zmena z dôvodu menej presných alebo chýbajúcich údajov než aká bola použitá v predchádzajúcom období
nc = žiadna zmena (napr. celkový stav vyhodnocovaný iba v roku 2013, ale predpokladá sa, že to isté hodnotenie by mal aj v roku 2007 ak by bol vtedy hodnotený)
Biotopy reportované Slovenskou republikou a ich stav v porovaní dvoch reportovacíh periód
|
Skupina |
Názov (EN) |
Kód |
Rok |
ALP |
PAN |
|
Lesné biotopy |
Acidophilous Picea forests of the montane to alpine levels (Vaccinio-Piceetea) |
9410 |
2013 |
U1= U1
|
|
|
|
Alluvial forests with Alnus glutinosa and Fraxinus excelsior (Alno-Padion, Alnion incanae, Salicion albae) |
91E0 |
2013 |
U1= U1
|
U2- U2-
|
|
|
Alpine Larix decidua and/or Pinus cembra forests |
9420 |
2013 |
FV FV
|
|
|
|
Asperulo-Fagetum beech forests |
9130 |
2013 |
FV FV
|
FV FV
|
|
|
Bog woodland |
91D0 |
2013 |
U1- U1-
|
|
|
|
Central European lichen Scots pine forestsT |
91T0 |
2013 |
|
U2-
nc |
|
|
Euro-Siberian steppic woods with Quercus spp. |
91I0 |
2013 |
FV FV
|
U2= U2
|
|
|
Galio-Carpinetum oak-hornbeam forests |
9170 |
2013 |
U2- U2-
|
|
|
|
Luzulo-Fagetum beech forests |
9110 |
2013 |
FV FV
|
U1= U1
|
|
|
Medio-European limestone beech forests of the Cephalanthero-Fagion |
9150 |
2013 |
FV FV
|
FV FV
|
|
|
Medio-European subalpine beech woods with Acer and Rumex arifolius |
9140 |
2013 |
FV FV
|
|
|
|
Old acidophilous oak woods with Quercus robur on sandy plains |
9190 |
2013 |
|
U1= U1
|
|
|
Pannonian woods with Quercus pubescens |
91H0 |
2013 |
U1= U1
|
FV FV
|
|
|
Pannonian-Balkanic turkey oak –sessile oak forests |
91M0 |
2013 |
U1= U1
|
U1= U1
|
|
|
Pannonic inland sand dune thicket (Junipero-Populetum albae) |
91N0 |
2013 |
|
U2- U2-
|
|
|
Pannonic woods with Quercus petraea and Carpinus betulus |
91G0 |
2013 |
FV FV
|
U1= U1
|
|
|
Riparian mixed forests of Quercus robur, Ulmus laevis and Ulmus minor, Fraxinus excelsior or Fraxinus angustifolia, along the great rivers (Ulmenion minoris) |
91F0 |
2013 |
FV FV
|
U2= U2
|
|
|
Tilio-Acerion forests of slopes, screes and ravines |
9180 |
2013 |
U1= U1
|
U1= U1
|
|
|
Western Carpathian calcicolous Pinus sylvestris forests |
91Q0 |
2013 |
FV FV
|
|
|
Skalné biotopy |
Calcareous and calcshist screes of the montane to alpine levels (Thlaspietea rotundifolii) |
8120 |
2013 |
FV FV
|
|
|
|
Calcareous rocky slopes with chasmophytic vegetation |
8210 |
2013 |
FV FV
|
FV FV
|
|
|
Caves not open to the public |
8310 |
2013 |
XX XX
|
XX XX
|
|
|
Medio-European calcareous scree of hill and montane levels |
8160 |
2013 |
FV FV
|
FV FV
|
|
|
Medio-European upland siliceous screes |
8150 |
2013 |
FV FV
|
FV FV
|
|
|
Siliceous rock with pioneer vegetation of the Sedo-Scleranthion or of the Sedo albi-Veronicion dillenii |
8230 |
2013 |
XX XX
|
XX XX
|
|
|
Siliceous rocky slopes with chasmophytic vegetation |
8220 |
2013 |
FV FV
|
FV FV
|
|
|
Siliceous scree of the montane to snow levels (Androsacetalia alpinae and Galeopsietalia ladani) |
8110 |
2013 |
FV FV
|
|
|
Slatiny a rašeliniská |
Active raised bogs |
7110 |
2013 |
U1= U1
|
|
|
|
Alkaline fens |
7230 |
2013 |
U1- U1 a |
U2= U1 a |
|
|
Calcareous fens with Cladium mariscus and species of the Caricion davallianae |
7210 |
2013 |
U1= U1
|
|
|
|
Degraded raised bogs still capable of natural regeneration |
7120 |
2013 |
U1= U1
|
|
|
|
Petrifying springs with tufa formation (Cratoneurion) |
7220 |
2013 |
U1= U1
|
|
|
|
Transition mires and quaking bogs |
7140 |
2013 |
U1= U1
|
U1= U1
|
|
Travinno bylinné biotopy |
Alluvial meadows of river valleys of the Cnidion dubii |
6440 |
2013 |
|
U1= U1
|
|
|
Alpine and subalpine calcareous grasslands |
6170 |
2013 |
U1= U1
|
|
|
|
Hydrophilous tall herb fringe communities of plains and of the montane to alpine levels |
6430 |
2013 |
FV U1 b1 |
FV U1 b1 |
|
|
Lowland hay meadows (Alopecurus pratensis, Sanguisorba officinalis) |
6510 |
2013 |
FV FV
|
U1= U1
|
|
|
Molinia meadows on calcareous, peaty or clayey-silt-laden soils (Molinion caeruleae) |
6410 |
2013 |
U1= U1
|
U1= U1
|
|
|
Mountain hay meadows |
6520 |
2013 |
U1= U1
|
|
|
|
Pannonic loess steppic grasslands |
6250 |
2013 |
|
U1- U1 b1 |
|
|
Pannonic sand steppes |
6260 |
2013 |
|
U2- U2-
|
|
|
Rupicolous calcareous or basophilic grasslands of the Alysso-Sedion albi |
6110 |
2013 |
FV U1 b1 |
FV U1 b1 |
|
|
Rupicolous pannonic grasslands (Stipo-Festucetalia pallentis) |
6190 |
2013 |
FV FV
|
FV FV
|
|
|
Semi-natural dry grasslands and scrubland facies on calcareous substrates (Festuco-Brometalia) (* important orchid sites) |
6210 |
2013 |
U1= U1
|
U1= XX b1 |
|
|
Siliceous alpine and boreal grasslands |
6150 |
2013 |
U1= U1
|
|
|
|
Species-rich Nardus grasslands, on silicious substrates in mountain areas (and submountain areas in Continental Europe) |
6230 |
2013 |
U1- U1 b1 |
|
|
|
Sub-Pannonic steppic grasslands |
6240 |
2013 |
U1- U1 b1 |
U1- U1 b1 |
|
|
Xeric sand calcareous grasslands |
6120 |
2013 |
|
U2- U2-
|
|
Kroviny |
Juniperus communis formations on heaths or calcareous grasslands |
5130 |
2013 |
U1= FV b1 |
U1= U1
|
|
Vresoviská a kríčkové biotopy |
Alpine and Boreal heaths |
4060 |
2013 |
FV FV
|
|
|
|
Bushes with Pinus mugo and Rhododendron hirsutum (Mugo-Rhododendretum hirsuti) |
4070 |
2013 |
FV FV
|
|
|
|
European dry heaths |
4030 |
2013 |
FV FV
|
FV FV
|
|
|
Sub-Arctic Salix spp. scrub |
4080 |
2013 |
U1= U1
|
|
|
|
Subcontinental peri-Pannonic scrub |
40A0 |
2013 |
U1= U1
|
U1= U1
|
|
Vodné biotopy |
Alpine rivers and the herbaceous vegetation along their banks |
3220 |
2013 |
FV U1 b1 |
|
|
|
Alpine rivers and their ligneous vegetation with Myricaria germanica |
3230 |
2013 |
FV U1 b1 |
|
|
|
Alpine rivers and their ligneous vegetation with Salix elaeagnos |
3240 |
2013 |
U1= U1
|
|
|
|
Hard oligo-mesotrophic waters with benthic vegetation of Chara spp. |
3140 |
2013 |
U1- XX b1 |
U1- XX b1 |
|
|
Natural dystrophic lakes and ponds |
3160 |
2013 |
U1= U1
|
U1= U1
|
|
|
Natural eutrophic lakes with Magnopotamion or Hydrocharition — type vegetation |
3150 |
2013 |
FV XX b1 |
U1= U1
|
|
|
Oligotrophic to mesotrophic standing waters with vegetation of the Littorelletea uniflorae and/or of the Isoëto-Nanojuncetea |
3130 |
2013 |
FV U1 b1 |
U1= U1
|
|
|
Rivers with muddy banks with Chenopodion rubri p.p. and Bidention p.p. vegetation |
3270 |
2013 |
XX U2 e |
XX U2 e |
|
|
Water courses of plain to montane levels with the Ranunculion fluitantis and Callitricho-Batrachion vegetation |
3260 |
2013 |
FV XX b1 |
FV XX b1 |
|
Pieskové biotopy |
Pannonic inland dunes |
2340 |
2013 |
|
U1= U1
|
|
Slaniská |
Inland salt meadows |
1340 |
2013 |
U2= U2
|
U2+ U2 a |
|
|
Pannonic salt steppes and salt marshes |
1530 |
2013 |
|
U2= U2
|
Druhy reportované Slovenskou republikou a ich stav v porovaní dvoch reportovacíh periód
|
Skupina |
Názov taxónu |
Kód |
Rok |
ALP |
PAN |
|
Nižšie rastliny |
Buxbaumia viridis |
1386 |
2013 |
U1= XX b1 |
|
|
|
Dicranum viride |
1381 |
2013 |
XX XX
|
|
|
|
Drepanocladus vernicosus |
1393 |
2013 |
U1= XX b1 |
|
|
|
Leucobryum glaucum |
1400 |
2013 |
U1+ U1+
|
U1+ U1+
|
|
|
Mannia triandra |
1379 |
2013 |
XX XX
|
|
|
|
Ochyraea tatrensis |
4119 |
2013 |
XX XX
|
|
|
|
Scapania massalongii |
1394 |
2013 |
XX XX
|
|
|
|
Sphagnum spp. |
1409 |
2013 |
U1+ U1+
|
U1= U1
|
|
|
Tortella rigens |
1988 |
2013 |
XX XX
|
|
|
Vyššie rastliny |
Aconitum firmum ssp. moravicum |
4109 |
2013 |
U1= U1
|
|
|
|
Adenophora lilifolia |
4068 |
2013 |
FV XX b1 |
U1= FV b1 |
|
|
Angelica palustris |
1617 |
2013 |
|
U2- U2 b1 |
|
|
Apium repens |
1614 |
2013 |
|
U2= U2
|
|
|
Artemisia eriantha |
1763 |
2013 |
FV FV
|
|
|
|
Asplenium adulterinum |
4066 |
2013 |
U1= FV a |
|
|
|
Campanula serrata |
4070 |
2013 |
FV FV
|
|
|
|
Cirsium brachycephalum |
4081 |
2013 |
|
U1= U1
|
|
|
Cochlearia tatrae |
4090 |
2013 |
FV FV
|
|
|
|
Colchicum arenarium |
2285 |
2013 |
|
U1= U2 a |
|
|
Crambe tataria |
4091 |
2013 |
|
FV FV
|
|
|
Cyclamen fatrense |
4107 |
2013 |
FV FV
|
|
|
|
Cypripedium calceolus |
1902 |
2013 |
U1= U1
|
U1= U1
|
|
|
Daphne arbuscula |
2159 |
2013 |
FV FV
|
|
|
|
Dianthus lumnitzeri |
4075 |
2013 |
U1= U1
|
U2= U2
|
|
|
Dianthus nitidus |
2074 |
2013 |
U1= U1
|
|
|
|
Dracocephalum austriacum |
1689 |
2013 |
U2- U2-
|
U1= U1
|
|
|
Echium russicum |
4067 |
2013 |
U2= U2
|
U2- U2-
|
|
|
Eleocharis carniolica |
1898 |
2013 |
U2= U2
|
|
|
|
Ferula sadleriana |
2170 |
2013 |
U1= U1
|
|
|
|
Galanthus nivalis |
1866 |
2013 |
FV FV
|
FV FV
|
|
|
Gladiolus palustris |
4096 |
2013 |
|
U2= U2
|
|
|
Himantoglossum adriaticum |
4104 |
2013 |
U2= U2
|
U2= U2
|
|
|
Himantoglossum caprinum |
2327 |
2013 |
U2= U2
|
U2= U2
|
|
|
Iris aphylla ssp. hungarica |
4097 |
2013 |
U1= U1
|
U1= U1
|
|
|
Iris humilis ssp. arenaria |
4098 |
2013 |
|
U2= U2
|
|
|
Ligularia sibirica |
1758 |
2013 |
U1= U1
|
|
|
|
Lindernia procumbens |
1725 |
2013 |
|
U1= XX b1 |
|
|
Liparis loeselii |
1903 |
2013 |
U1= U2 b1 |
U2= U2
|
|
|
Lycopodium spp. |
1413 |
2013 |
FV FV
|
U1+ U1+
|
|
|
Marsilea quadrifolia |
1428 |
2013 |
|
U2- U2-
|
|
|
Onosma tornensis |
2203 |
2013 |
U1- U1-
|
U1= U1
|
|
|
Pulsatilla grandis |
2093 |
2013 |
U1- U1-
|
U1= U1
|
|
|
Pulsatilla patens |
1477 |
2013 |
U1= U1
|
U2- FV b1 |
|
|
Pulsatilla pratensis ssp. hungarica |
4110 |
2013 |
|
U2- U2-
|
|
|
Pulsatilla slavica |
2094 |
2013 |
U1= U1
|
|
|
|
Pulsatilla subslavica |
4111 |
2013 |
U1= U1
|
|
|
|
Serratula lycopifolia |
4087 |
2013 |
U2= U2
|
|
|
|
Tephroseris longifolia ssp. moravica |
4088 |
2013 |
U1= U1
|
|
|
|
Thlaspi jankae |
2120 |
2013 |
U1= U1
|
U1= U1
|
|
|
Tozzia carpathica |
4116 |
2013 |
FV FV
|
|
|
Mäkkýše |
Anisus vorticulus |
4056 |
2013 |
|
U1- XX b1 |
|
|
Helix pomatia |
1026 |
2013 |
FV FV
|
FV FV
|
|
|
Sadleriana pannonica |
4063 |
2013 |
U1= U2 b1 |
U1= U2 b1 |
|
|
Unio crassus |
1032 |
2013 |
FV FV
|
U1= U2- b1 |
|
|
Vertigo angustior |
1014 |
2013 |
FV FV
|
FV U1- b1 |
|
|
Vertigo geyeri |
1013 |
2013 |
U1= XX b1 |
|
|
|
Vertigo moulinsiana |
1016 |
2013 |
U1= U1
|
U2- XX b1 |
|
Článkonožce |
Astacus astacus |
1091 |
2013 |
U1= U1
|
XX XX
|
|
|
Austropotamobius torrentium |
1093 |
2013 |
U2= U2
|
|
|
|
Bolbelasmus unicornis |
4011 |
2013 |
U2= U2
|
U2= U2
|
|
|
Boros schneideri |
1920 |
2013 |
U2- XX b1 |
|
|
|
Callimorpha quadripunctaria |
1078 |
2013 |
FV FV
|
FV FV
|
|
|
Carabus hungaricus |
4013 |
2013 |
|
U2= U2
|
|
|
Carabus variolosus |
4014 |
2013 |
XX XX
|
XX XX
|
|
|
Carabus zawadzkii |
4015 |
2013 |
XX XX
|
|
|
|
Cerambyx cerdo |
1088 |
2013 |
U1- XX b1 |
U1- XX b1 |
|
|
Coenagrion ornatum |
4045 |
2013 |
U1= U2+ b1 |
U2= U2
|
|
|
Coenonympha hero |
1070 |
2013 |
FV FV
|
|
|
|
Colias myrmidone |
4030 |
2013 |
U2- U2-
|
U2- U2-
|
|
|
Cordulegaster heros |
4046 |
2013 |
FV U2 b1 |
U1= U2 b1 |
|
|
Cucujus cinnaberinus |
1086 |
2013 |
FV
|
FV XX b1 |
|
|
Dioszeghyana schmidtii |
4032 |
2013 |
|
FV FV
|
|
|
Duvalius hungaricus |
4018 |
2013 |
FV FV
|
FV FV
|
|
|
Eriogaster catax |
1074 |
2013 |
FV FV
|
FV FV
|
|
|
Graphoderus bilineatus |
1082 |
2013 |
U2- U2-
|
U2- U2-
|
|
|
Hypodryas maturna |
1052 |
2013 |
U1= U1
|
FV FV
|
|
|
Isophya stysi |
4050 |
2013 |
FV U1 b1 |
FV U1 b1 |
|
|
Leptidea morsei |
4036 |
2013 |
FV U2 b1 |
|
|
|
Leucorrhinia pectoralis |
1042 |
2013 |
U2= U2
|
FV U1- a |
|
|
Limoniscus violaceus |
1079 |
2013 |
U1- XX b1 |
U2- U2+ b1 |
|
|
Lopinga achine |
1067 |
2013 |
U2= U2
|
U2= U2
|
|
|
Lucanus cervus |
1083 |
2013 |
FV FV
|
FV FV
|
|
|
Lycaena dispar |
1060 |
2013 |
FV FV
|
FV FV
|
|
|
Maculinea arion |
1058 |
2013 |
U1- U1-
|
U2- U2-
|
|
|
Maculinea nausithous |
1061 |
2013 |
FV U2 b1 |
FV U1 b1 |
|
|
Maculinea teleius |
1059 |
2013 |
FV FV
|
FV FV
|
|
|
Odontopodisma rubripes |
4052 |
2013 |
FV XX b1 |
FV XX b1 |
|
|
Ophiogomphus cecilia |
1037 |
2013 |
U1= U2 b1 |
U1- U1-
|
|
|
Osmoderma eremita |
1084 |
2013 |
U1- XX b1 |
U1- XX b1 |
|
|
Paracaloptenus caloptenoides |
4053 |
2013 |
XX XX
|
U1x
nc |
|
|
Parnassius apollo |
1057 |
2013 |
U1- U1-
|
|
|
|
Parnassius mnemosyne |
1056 |
2013 |
FV FV
|
FV FV
|
|
|
Pholidoptera transsylvanica |
4054 |
2013 |
FV U1 b1 |
FV
|
|
|
Proserpinus proserpina |
1076 |
2013 |
XX XX
|
XX XX
|
|
|
Pseudogaurotina excellens |
4024 |
2013 |
XX XX
|
|
|
|
Rhysodes sulcatus |
4026 |
2013 |
U1- XX b1 |
U2- XX b1 |
|
|
Rosalia alpina |
1087 |
2013 |
FV FV
|
FV FV
|
|
|
Saga pedo |
1050 |
2013 |
U1= U1
|
U1= U1
|
|
|
Stenobothrus eurasius |
4055 |
2013 |
FV U1- b1 |
U1= U1
|
|
|
Stylurus flavipes |
1040 |
2013 |
|
FV U2 b1 |
|
|
Zerynthia polyxena |
1053 |
2013 |
FV FV
|
FV FV
|
|
Ryby |
Aspius aspius |
1130 |
2013 |
FV FV
|
FV FV
|
|
|
Barbus barbus |
5085 |
2013 |
U1- U1-
|
U1- FV a |
|
|
Barbus meridionalis |
1138 |
2013 |
U1- XX b1 |
U1- XX b1 |
|
|
Cobitis taenia |
1149 |
2013 |
XX XX
|
U1- XX b1 |
|
|
Cottus gobio |
1163 |
2013 |
FV XX b1 |
U2- U1- a |
|
|
Eudontomyzon danfordi |
4123 |
2013 |
U1- XX
|
U1- XX
|
|
|
Eudontomyzon mariae |
2484 |
2013 |
XX XX
|
XX XX
|
|
|
Gobio kessleri |
2511 |
2013 |
U1- XX b1 |
U1- XX b1 |
|
|
Gobio uranoscopus |
1122 |
2013 |
U1- XX b1 |
U1- XX b1 |
|
|
Gobio vladykovi |
6158 |
2013 |
XX
|
U1-
b1 |
|
|
Gymnocephalus baloni |
2555 |
2013 |
|
U2- XX b1 |
|
|
Gymnocephalus schraetzer |
1157 |
2013 |
U2- XX b1 |
U2- XX b1 |
|
|
Hucho hucho |
1105 |
2013 |
XX XX
|
U1- XX b1 |
|
|
Lampetra planeri |
1096 |
2013 |
U1x XX b1 |
|
|
|
Misgurnus fossilis |
1145 |
2013 |
|
U1- XX b1 |
|
|
Pelecus cultratus |
2522 |
2013 |
|
XX XX
|
|
|
Rhodeus sericeus amarus |
1134 |
2013 |
FV FV
|
FV FV
|
|
|
Rutilus virgo |
5345 |
2013 |
XX XX
|
U1- XX b1 |
|
|
Sabanejewia aurata |
1146 |
2013 |
XX XX
|
U1- XX b1 |
|
|
Thymallus thymallus |
1109 |
2013 |
U2- U1 a |
|
|
|
Umbra krameri |
2011 |
2013 |
|
U2- XX b1 |
|
|
Zingel streber |
1160 |
2013 |
XX XX
|
U1x XX b1 |
|
|
Zingel zingel |
1159 |
2013 |
|
XX XX
|
|
Obojživelníky |
Bombina bombina |
1188 |
2013 |
U2= U2
|
U1= U1
|
|
|
Bombina variegata |
1193 |
2013 |
U1- U1-
|
XX XX
|
|
|
Bufo viridis |
1201 |
2013 |
U1+ U1+
|
U1+ U1+
|
|
|
Hyla arborea |
1203 |
2013 |
U1= U1
|
U1= U1
|
|
|
Pelobates fuscus |
1197 |
2013 |
U1- U1-
|
U1= U1
|
|
|
Rana arvalis |
1214 |
2013 |
|
U2- U2-
|
|
|
Rana dalmatina |
1209 |
2013 |
U1= U1
|
U1= U1
|
|
|
Rana esculenta |
1210 |
2013 |
U1= U1
|
U1- U1-
|
|
|
Rana lessonae |
1207 |
2013 |
U2- U2-
|
U2= U2
|
|
|
Rana ridibunda |
1212 |
2013 |
U1= U1
|
U1= U1
|
|
|
Rana temporaria |
1213 |
2013 |
FV FV
|
U1= U1
|
|
|
Triturus cristatus |
1166 |
2013 |
U2- U2-
|
|
|
|
Triturus dobrogicus |
1993 |
2013 |
U2- U2 b1 |
U1- U1-
|
|
|
Triturus montandoni |
2001 |
2013 |
U1- U1-
|
|
|
Plazy |
Ablepharus kitaibelii |
1276 |
2013 |
|
U1- U1 b1 |
|
|
Coronella austriaca |
1283 |
2013 |
U2- U1 a |
U2- U1 a |
|
|
Elaphe longissima |
1281 |
2013 |
U2= U1 b1 |
U2= U1 b1 |
|
|
Emys orbicularis |
1220 |
2013 |
|
U2- U2-
|
|
|
Lacerta agilis |
1261 |
2013 |
U1- FV a |
U1- FV a |
|
|
Lacerta viridis |
1263 |
2013 |
U1- U1-
|
U1- U1-
|
|
|
Lacerta vivipara pannonica |
5037 |
2013 |
|
U1- U1-
|
|
|
Natrix tessellata |
1292 |
2013 |
U1- U1-
|
U1- U1-
|
|
|
Podarcis muralis |
1256 |
2013 |
U1= FV a |
U1= FV a |
|
Cicavce |
Barbastella barbastellus |
1308 |
2013 |
XX XX
|
XX XX
|
|
|
Bison bonasus |
2647 |
2013 |
U1= U2+ a |
|
|
|
Canis lupus |
1352 |
2013 |
FV FV
|
U1= U1
|
|
|
Castor fiber |
1337 |
2013 |
FV FV
|
FV FV
|
|
|
Cricetus cricetus |
1339 |
2013 |
U1= U1
|
U1+ U1+
|
|
|
Dryomys nitedula |
1342 |
2013 |
U1= U1
|
U2+ U2+
|
|
|
Eptesicus nilssonii |
1313 |
2013 |
XX XX
|
|
|
|
Eptesicus serotinus |
1327 |
2013 |
XX XX
|
XX XX
|
|
|
Felis silvestris |
1363 |
2013 |
U1= U1
|
U1= U1
|
|
|
Lutra lutra |
1355 |
2013 |
U1+ U1+
|
U1= U1
|
|
|
Lynx lynx |
1361 |
2013 |
U1= U1
|
U1= U1
|
|
|
Marmota marmota latirostris |
4003 |
2013 |
U2= U2
|
|
|
|
Martes martes |
1357 |
2013 |
U1= U1
|
U1= U1
|
|
|
Microtus oeconomus mehelyi |
4004 |
2013 |
|
U2- U2-
|
|
|
Microtus tatricus |
2612 |
2013 |
U2= U2
|
|
|
|
Miniopterus schreibersii |
1310 |
2013 |
U2- U2-
|
U2- U2-
|
|
|
Muscardinus avellanarius |
1341 |
2013 |
U1= U1
|
U1= U1
|
|
|
Mustela eversmanii |
2633 |
2013 |
|
U2= U2
|
|
|
Mustela putorius |
1358 |
2013 |
U1- U1-
|
U1- U1-
|
|
|
Myotis bechsteinii |
1323 |
2013 |
XX XX
|
XX XX
|
|
|
Myotis blythii |
1307 |
2013 |
XX XX
|
XX XX
|
|
|
Myotis brandtii |
1320 |
2013 |
XX XX
|
XX XX
|
|
|
Myotis dasycneme |
1318 |
2013 |
XX XX
|
XX XX
|
|
|
Myotis daubentonii |
1314 |
2013 |
XX XX
|
XX XX
|
|
|
Myotis emarginatus |
1321 |
2013 |
XX XX
|
XX XX
|
|
|
Myotis myotis |
1324 |
2013 |
U1- U1-
|
U1- U1-
|
|
|
Myotis mystacinus |
1330 |
2013 |
XX XX
|
XX XX
|
|
|
Myotis nattereri |
1322 |
2013 |
XX XX
|
U1- U1-
|
|
|
Nyctalus lasiopterus |
1328 |
2013 |
XX XX
|
XX XX
|
|
|
Nyctalus leisleri |
1331 |
2013 |
XX XX
|
XX XX
|
|
|
Nyctalus noctula |
1312 |
2013 |
U2- U2-
|
U2- U2-
|
|
|
Pipistrellus nathusii |
1317 |
2013 |
XX XX
|
XX XX
|
|
|
Pipistrellus pipistrellus |
1309 |
2013 |
XX XX
|
XX XX
|
|
|
Pipistrellus pygmaeus |
5009 |
2013 |
XX XX
|
XX XX
|
|
|
Plecotus auritus |
1326 |
2013 |
XX XX
|
XX XX
|
|
|
Plecotus austriacus |
1329 |
2013 |
XX XX
|
XX XX
|
|
|
Rhinolophus euryale |
1305 |
2013 |
U1- U1-
|
U1= U1
|
|
|
Rhinolophus ferrumequinum |
1304 |
2013 |
U1- U1-
|
U1- U1-
|
|
|
Rhinolophus hipposideros |
1303 |
2013 |
U1- U1-
|
U1- U1-
|
|
|
Rupicapra rupicapra tatrica |
4006 |
2013 |
U1+ U2+ a |
|
|
|
Sicista betulina |
1343 |
2013 |
U1- U1-
|
|
|
|
Spermophilus citellus |
1335 |
2013 |
U2- U2-
|
U2- U2-
|
|
|
Ursus arctos |
1354 |
2013 |
FV FV
|
|
|
|
Vespertilio murinus |
1332 |
2013 |
XX XX
|
XX XX
|
|
Iné bezstavovce |
Hirudo medicinalis |
1034 |
2013 |
|
FV U1- b1 |
V nasledujúcej tabuľke sú uvedené druhy označené ako príležitostný (OCC), nastupujúci (ARR), vyhynutý po tom, čo smernica o biotopoch nadobudla účinnosť (EX), zaniknutý pred tým, ako smernica o biotopoch nadobudla účinnosť (PEX), marginálny (Mar.),. Okrem toho sú to uvedené druhy, pre ktoré bol zaslaný dobrovoľný report (OP) a druhy málo poznané a majú vedeckú nedostatočnosť v poznaní (SR).
|
Skupina |
Názov taxónu |
Kód |
Rok |
ALP |
PAN |
|
Nižšie rastliny |
Meesia longiseta |
1389 |
2013 |
PEX
|
|
|
|
Orthotrichum rogeri |
1387 |
2013 |
PEX
|
|
|
Vyššie rastliny |
Aldrovanda vesiculosa |
1516 |
2013 |
|
PEX
|
|
|
Coleanthus subtilis |
1887 |
2013 |
PEX
|
|
|
|
Spiranthes aestivalis |
1900 |
2013 |
|
PEX
|
|
|
Thesium ebracteatum |
1437 |
2013 |
|
PEX
|
|
Mäkkýše |
Theodoxus transversalis |
4064 |
2013 |
|
PEX
|
|
Článknožce |
Austropotamobius torrentium |
1093 |
2013 |
|
PEX
|
|
|
Chondrosoma fiduciarium |
4029 |
2013 |
|
PEX
|
|
|
Coenagrion mercuriale |
1044 |
2013 |
|
PEX
|
|
|
Coenonympha oedippus |
1071 |
2013 |
|
PEX
|
|
|
Dytiscus latissimus |
1081 |
2013 |
|
PEX
|
|
|
Euphydryas aurinia |
1065 |
2013 |
|
PEX
|
|
|
Leptidea morsei |
4036 |
2013 |
|
SR XX
|
|
|
Leucorrhinia albifrons |
1038 |
2013 |
|
PEX
|
|
|
Lycaena helle |
4038 |
2013 |
PEX
|
PEX
|
|
|
Morimus funereus |
1089 |
2013 |
SR
|
SR
|
|
|
Nymphalis vaualbum |
4039 |
2013 |
PEX
|
PEX
|
|
|
Phryganophilus ruficollis |
4021 |
2013 |
SR
|
|
|
|
Polyommatus eroides |
4042 |
2013 |
PEX
|
PEX
|
|
|
Stephanopachys substriatus |
1927 |
2013 |
SR
|
|
|
Ryby |
Huso huso |
2489 |
2013 |
|
PEX
|
|
|
Salmo salar |
1106 |
2013 |
PEX
|
|
|
Obojživelníky |
Triturus cristatus |
1166 |
2013 |
|
SR
|
|
Cicavce |
Hypsugo savii |
5365 |
2013 |
|
OCC XX XX
|
|
|
Mustela eversmanii |
2633 |
2013 |
OCC U2= U2
|
|
|
|
Mustela lutreola |
1356 |
2013 |
PEX
|
|
|
|
Myotis alcathoe |
5003 |
2013 |
|
OCC XX XX
|
|
|
Pipistrellus kuhlii |
2016 |
2013 |
|
OCC XX XX
|